







Membranes are vital because they separate the cell from the outside world. They also separate compartments inside the cell to protect important processes and events.
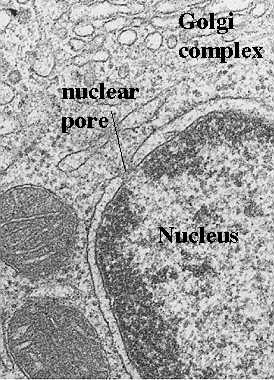
Cellular membranes have diverse functions in the different regions and organelles of a cell. However, at the electron microscopic level, they share a common structure following routine preparative steps. The above figure shows the typical “Unit” membrane. which resembles a railroad track with two dense lines separated by a clear space. This figure actually shows two adjacent plasma membranes, both of which have the "unit membrane” structure.
(Above figure taken from Bloom and Fawcett, A Textbook of Histology, Chapman and Hall, N.Y., 12th edition, 1994, Figure 1-2.)
To prepare for this part of the discussion on membranes, read pages 477-488 of your text
(Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y. Third edition, 1994.)

How did early cell biologists deduce membrane structure from electron microscopic images and the knowledge that membranes were lipoprotein complexes?
Note the material projecting from the plasma membrane both inside and outside of the cell. What do you think this is? Also, outside of the cell is in the upper half of photo. Can you find a vesicle with a unit membrane structure?
A historical perspective
In the early 1930's-40's, Danielli and Davson studied triglyceride lipid bilayers over a water surface. They found that they arranged themselves with the polar heads facing outward. However, they always formed droplets (oil in water) and the surface tension was much higher than that of cells. However, if you added proteins, the surface tension was reduced and the membranes flattened out.
Here is a diagram of their early model of the cell membrane.
What membrane functions might be allowed by this model?
In the 1950's Robertson noted the structure of membranes seen in the above electron micrographs. He saw no spaces for pores in the electron micrographs. He hypothesized that the railroad track appearance came from the binding of osmium tetroxide to proteins and polar groups of lipids.

What is missing in Robertson's model?
In 1966, Lenard and Singer noted that over 30% of membrane proteins were twisted into an alpha helix. This made it likely that there were many spherical proteins.
Could one build a Davson-Danielli or Robertson model with spherical proteins? What would happen to the size of the membrane?
Furthermore, what would happen if you unfolded the proteins, exposing non-polar (hydrophobic amino acid side chains) groups to the aqueous environment?
What kind of energy would the cell have to expend to keep proteins flattened in this state?
Singer studied phospholipid bilayers and found that they can form a flattened surface
on water, with no requirement for a protein coat. The turning point in the modeling
came with the advent of freeze fracture techniques.
This figure shows the inside of a membrane and the "bumps, grooves,
ridges". These were later found to be proteins. 
What conclusions could you draw about membrane structure from the photo to the left?
Basic Membrane Architecture
The purpose of this presentation will be to show how membranes are studied by the cell
biologist. First, we will look at basic unit membrane architecture. All membranes
contain proteins and lipid. However, the proportion of each varies depending on the
membrane. For example: Myelin, which insulates nerve fibers, contains only 18% protein
and 76% lipid. A electron micrograph of myelin is to the right.Mitochondrial inner membrane contain 76% protein and only 24%
lipid. Plasma membranes of human red blood cells and mouse liver contain
nearly equal amounts of proteins (44, 49% respectively) and lipids (43, 52% respectively).
Considering what you already know about these
cells or organelles, what would be the significance of these different proportions of
lipids and proteins?

As we said above, membrane architecture is that of a lipid bilayer. The lipids are amphipathic in that they have hydrophilic polar heads pointing out and the hydrophobic portion forming the core.

In addition to this figure, check out your text (Alberts et al, Molecular Biology of the Cell, Third edition, Garland Publishing, N.Y. 1994) on page 55 for a cartoon of a lipid bilayer formed by phospholipids and glycolipids. These figures are taken from your text, Figure 10-1, page 477. Proteins are embedded in the bilayer. They may pass through the bilayer (as transmembrane proteins), or they may be inserted at the cytoplasmic or exterior face.

A cross section of the bilayer is seen in this figure. As we will see in more detail below, the lipid molecules have a globular (polar) head and a straight region (non-polar). Each row of lipids is a leaflet. Therefore, the plasma membrane consists of two leaflets with the non-polar regions pointing inward.

Membrane Phospholipids
Lets take apart the membrane and examine each of its components. One of the principal types of lipid in the membrane include the phospholipids. These have a polar head group and two hydrocarbon tails. An example of a phospholipid is shown in this figure (right). The top region beginning with the NH3 is the polar group. It is connected by glycerol to two fatty acid tails. One of the tails is a straight chain fatty acid (saturated). The other has a kink in the tail because of a cis double bond (unsaturated). This kink influences packing and movement in the lateral plane of the membrane. Figure to the left is modified from
Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-10.
The figure below is from Wolfe S.L., Molecular and Cellular Biology, Wadsworth Publishing Company, 1993, p 155.

The figure in this paragraph shows how the phospholipids pack together in the two leaflets in the membrane. The presence of the cis double bond makes prevents tight packing and makes the bilayer difficult to freeze. Figure modified from

Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-7.
The lipid bilayer gives the membranes its fluid
characteristics. The following cartoon shows the effect of temperature on the
packing of the hydrocarbons. Note that a low temperatures, the bilayer is in a gel
state and tightly packed. At higher (body) temperatures, the bilayer actually
"melts' and the interior is fluid allowing the lipid molecules to move around,
rotate, exchange places. This also allows movement of other components of the
membrane. The figure below is from Wolfe S.L., Molecular and Cellular Biology,
Wadsworth Publishing Company, 1993.

Membrane Cholesterol
Another type of lipid in the membrane is cholesterol. The amount of cholesterol may vary with the type of membrane. Plasma membranes have nearly one cholesterol per phospholipid molecule. Other membranes (like those around bacteria) have no cholesterol. The following figure shows the steroid structure of cholesterol. The non-polar and polar regions are also illustrated in b) (Figure modified from
Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-8.

The cholesterol molecule inserts itself in the membrane with the same orientation as the phospholipid molecules. The figures show phospholipid molecules with a cholesterol molecule inbetween. Note that the polar head of the cholesterol is aligned with the polar head of the phospholipids. Figure modified from
Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-9; or
Wolfe S.L., Molecular and Cellular Biology, Wadsworth Publishing Company, 1993 (figure below).

Cholesterol molecules have several functions in the membrane:
- They immobilize the first few hydrocarbon groups of the phospholipid molecules. This makes the lipid bilayer less deformable and decreases its permeability to small water-soluble molecules. Without cholesterol (such as in a bacterium) a cell would need a cell wall.
- Cholesterol prevents crystallization of hydrocarbons and phase shifts in the membrane.
Return to Menu
Membrane Glycolipids

Glycolipids are also a constituent of membranes. In this figure, they are shown as blue sugar groups projecting into the extracellular space. They may microaggregate in the membrane. These components of the membrane may be protective, insulators, and sites of receptor binding. Among the molecules bound by glycososphingolipids include cell poisons such as cholera and tetanus toxins. The lower figure shows the chemical structure of two examples of glycososphingolipids.
Top Figure modified from Alberts et al. Molecular Biology of the Cell, Garland Publishing, N.Y., 1994, Third Edition, Figure 10-11.
Formation of "Microdomains"
Sphingolipids and cholesterol work together to help cluster proteins in a region called a "microdomain". They function as "rafts" or platforms for the attachment of proteins as membranes are moved around the cell and also during signal transduction. For more information about the rafts and "microdomains" see: Simons and Ikonen, Nature 387: 569, 1997. Also, R.E. Brown, J. Cell Science 111: 1-9, 1998
Membrane Proteins
As you study different organelles, you will learn about important membrane proteins that function for that particular organelle.
Transmembrane proteins are amphipathic, in that they have hydrophobic and hydrophilic regions that are oriented in the same regions in the lipid bilayer. Another name for them is "integral proteins". Other types of proteins may be linked only at the cytoplasmic surface (by attachment to a fatty acid chain), or at the external cell surface, attached by a oligosaccharide. Or, these non-transmembrane proteins may be bound to other membrane proteins. Collectively these are called "peripheral membrane proteins" .
We will be studying specific membrane proteins in later lectures (ion channels, proteins in endoplasmic reticulum, etc). Therefore, this presentation will not spend much time on them. Review pp 486 and 487 in your text for information on insertion of a transmembrane proteins. Proteins inserted once through the membrane are called "single-pass transmembrane proteins." Those that pass through several times are called "multipass transmembrane proteins". They form loops outside the membrane Later lecturers will spend more time on this as key proteins are introduced.
The following figure shows transmembrane proteins passing through the lipid bilayer. Can you find the multipass and single-pass transmembrane proteins in this figure? The figure below is from Wolfe S.L., Molecular and Cellular Biology, Wadsworth Publishing Company, 1993.
For more information, contact:
Gwen Childs, Ph.D.,FAAA
Professor and Chair
Department of Neurobiology and Developmental Sciences
University of Arkansas for Medical Sciences
Little Rock, AR 72205
For questions, contact this email address:
